Способ получения трансгенного растения ipomoea batatas с высоким накоплением каротиноида и антоциана и растение, полученное этим способом
Номер патента: 30683
Опубликовано: 15.12.2015
Авторы: ЛИ, Хенг-Сун, ПАК, Сунг Чул, КВОК, Санг-Су, КИМ, Сун ха, ЖОНГ, Жэ Чол









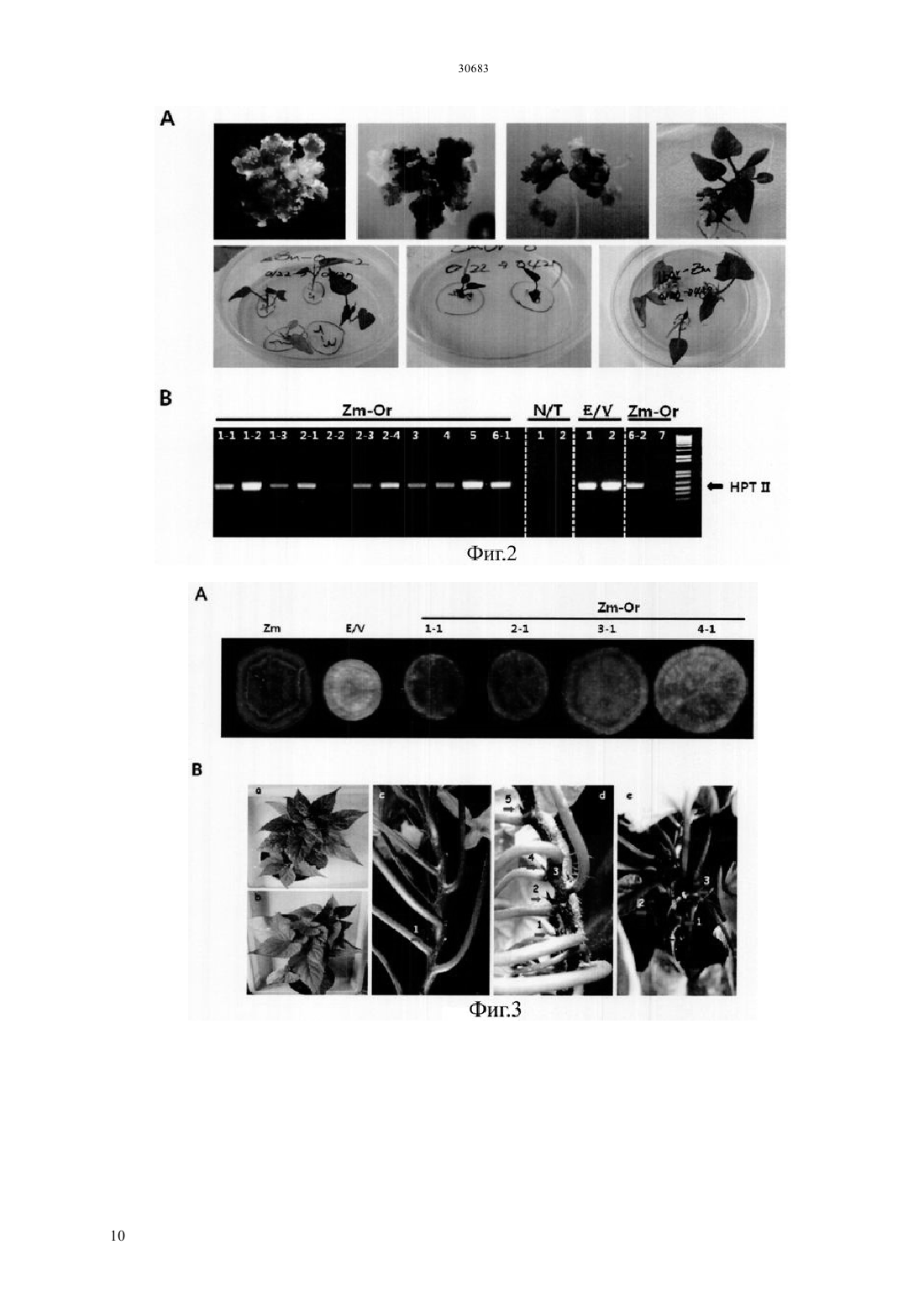
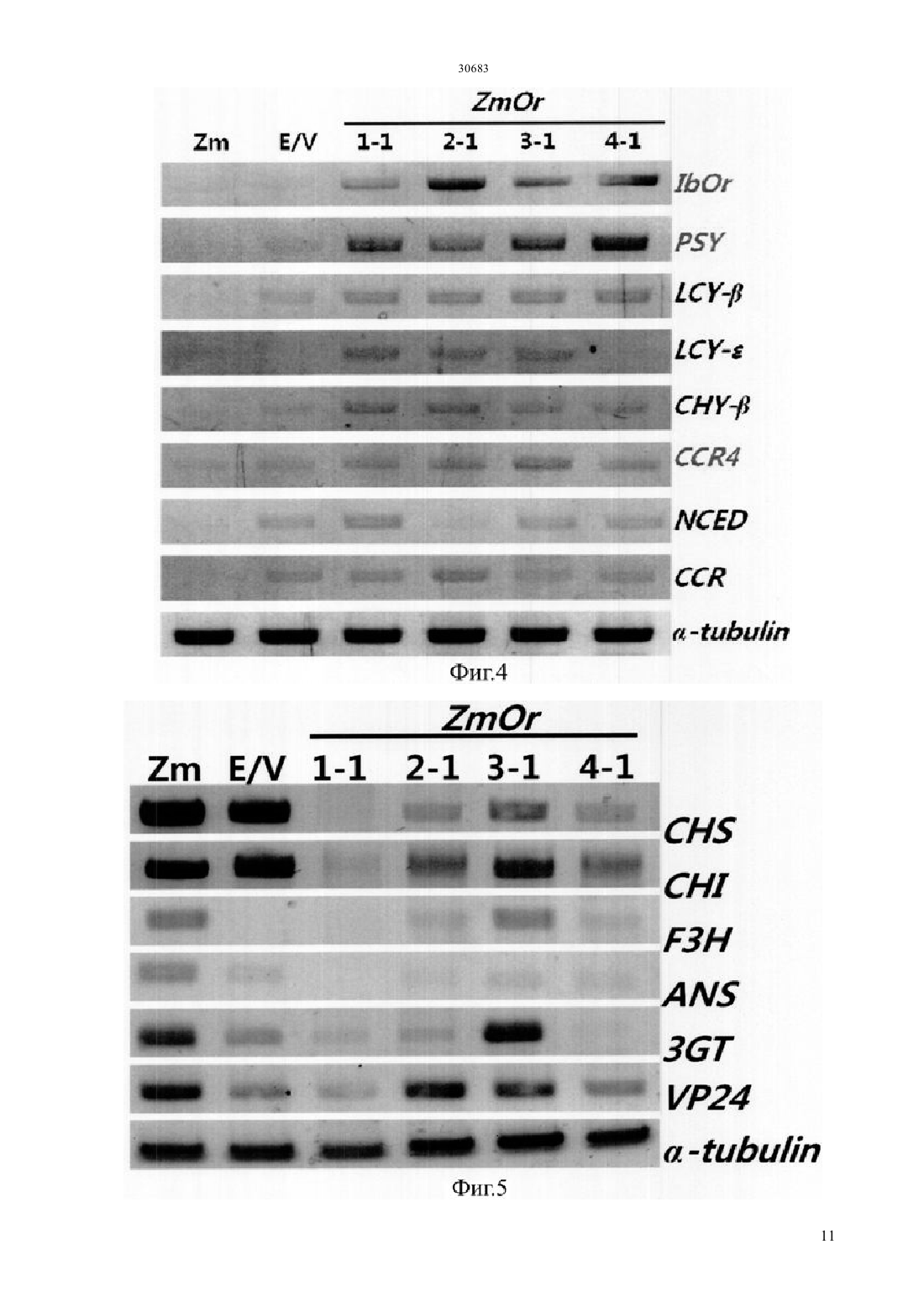
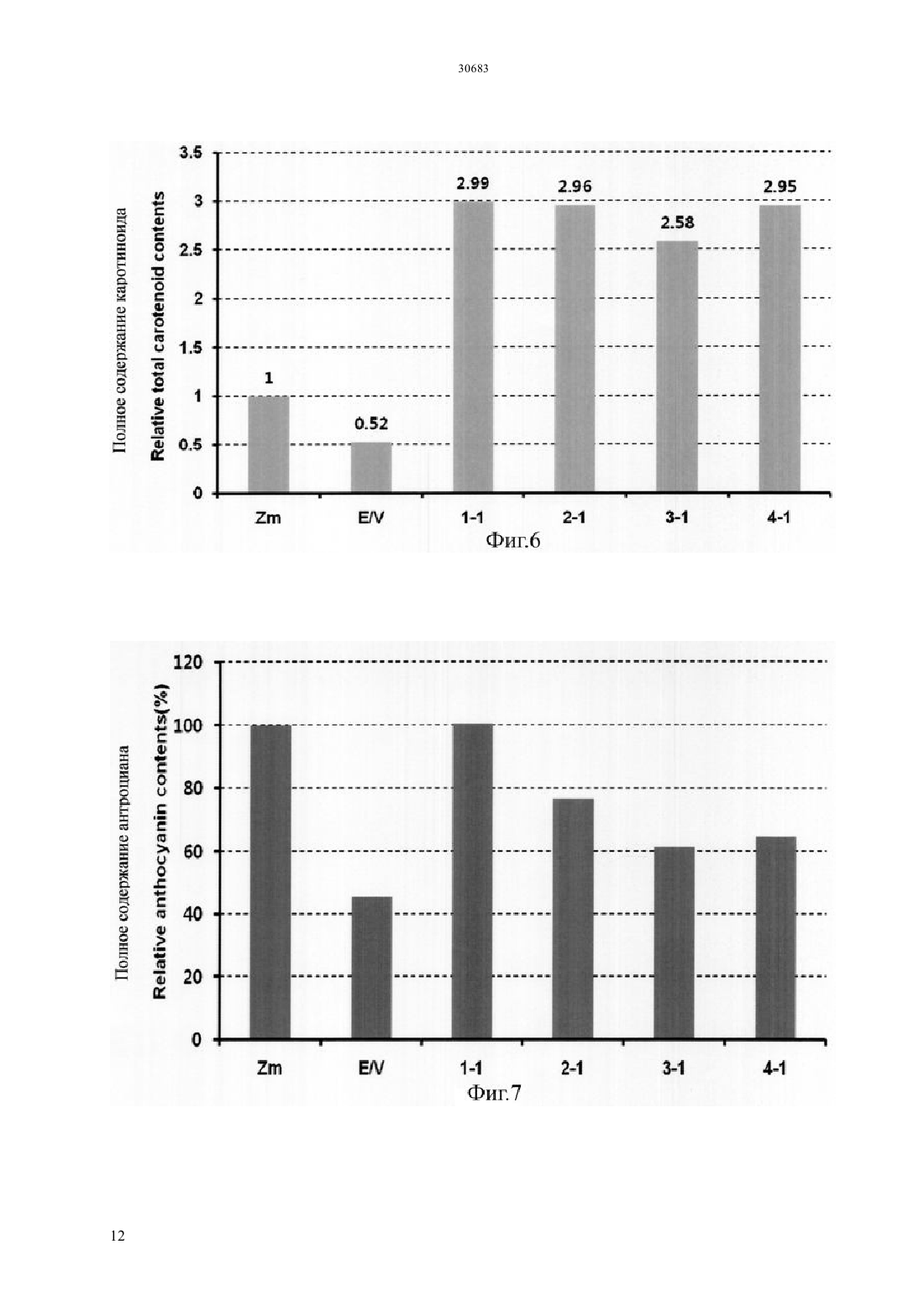
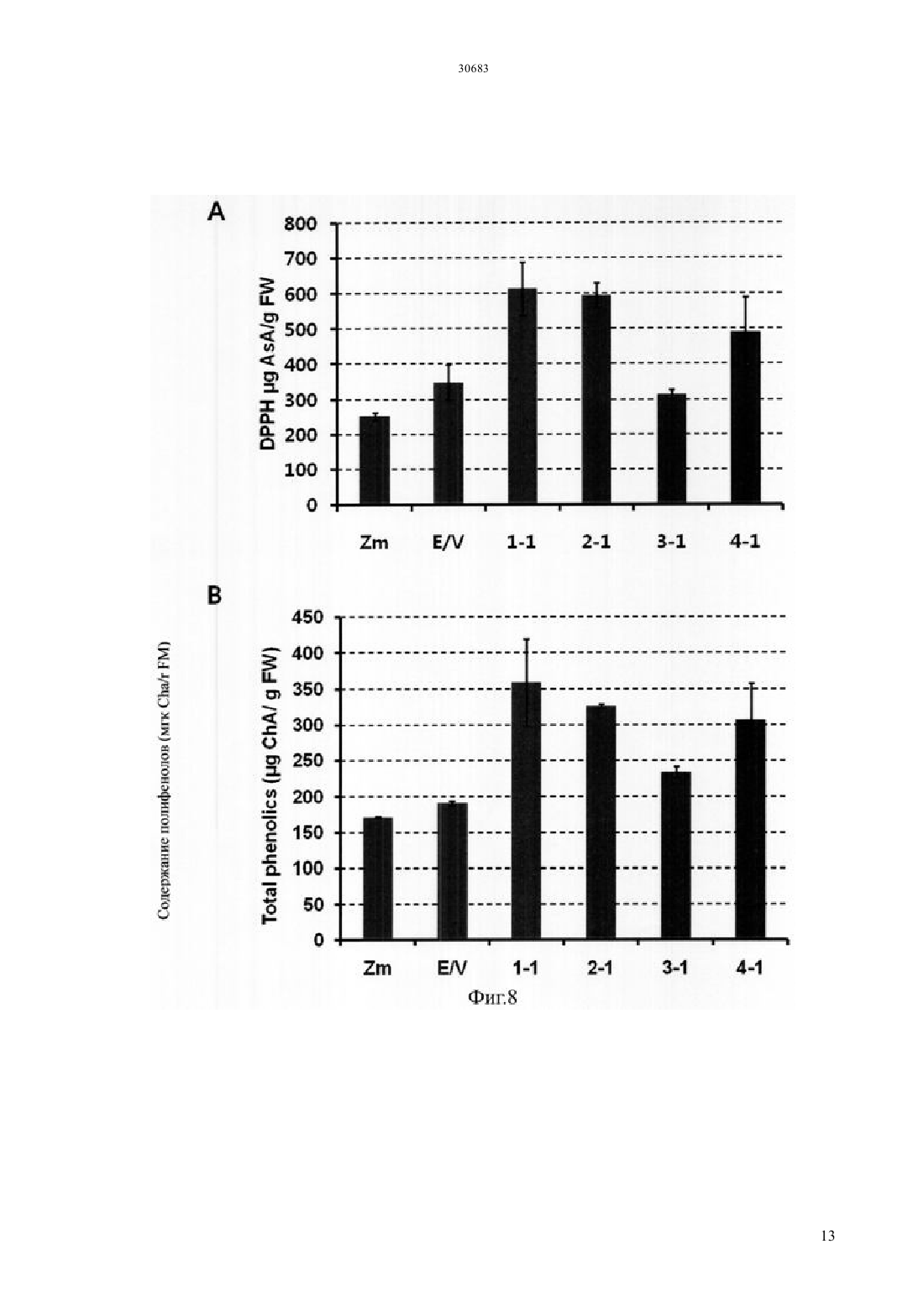
Текст
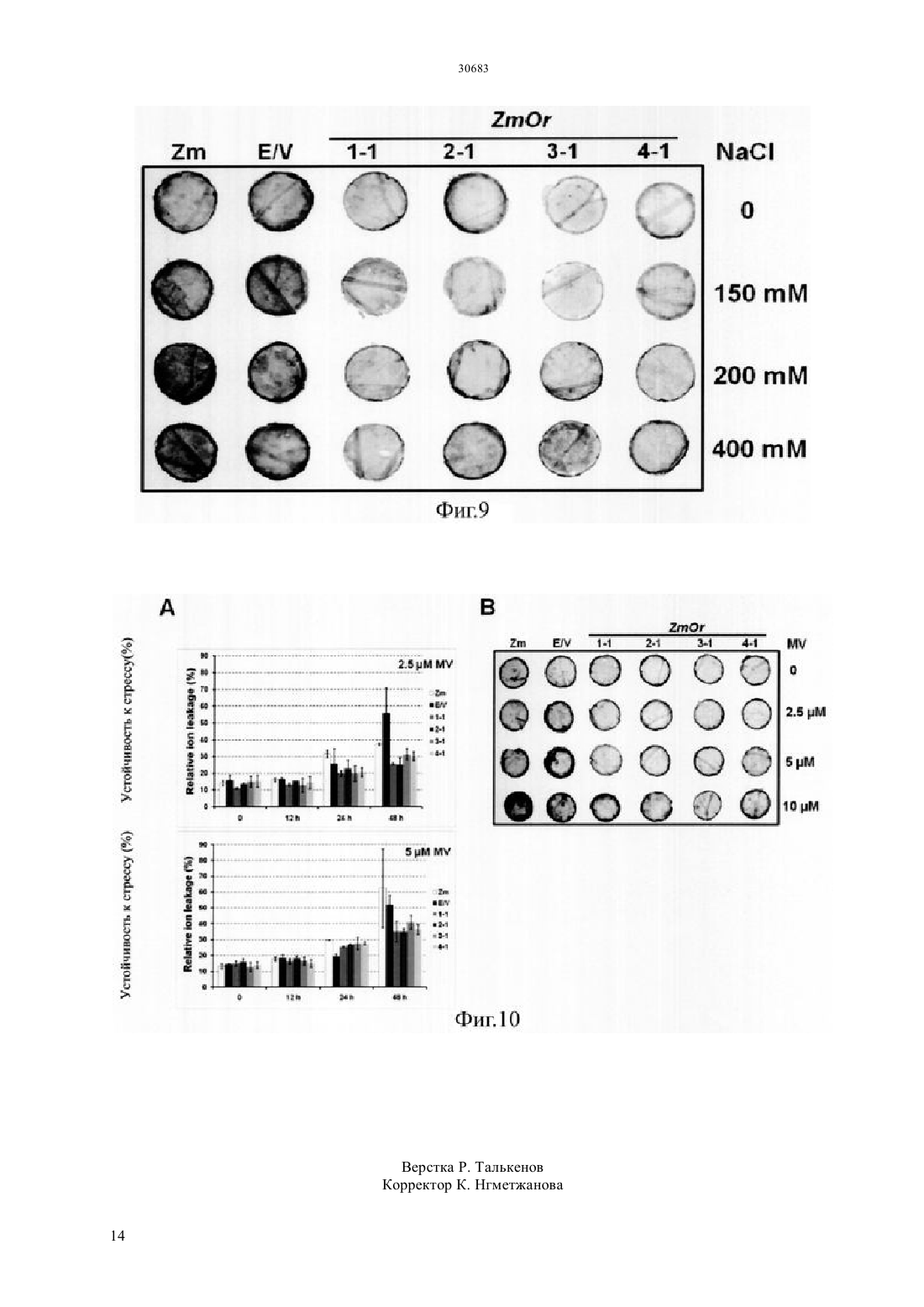
(51) 01 5/00 (2006.01) 12 15/29 (2006.01) 12 15/82 (2006.01) МИНИСТЕРСТВО ЮСТИЦИИ РЕСПУБЛИКИ КАЗАХСТАН ОПИСАНИЕ ИЗОБРЕТЕНИЯ К ПАТЕНТУ каротиноида и антоциана путем трансформации растения батата, способного накапливать антоциан,рекомбинантным вектором, содержащим модификацию гена, содержащую специфическую последовательность, искусственно вставленную в ген , полученный изк трансгенному растению батата с высоким накоплением каротиноида и антоциана, которое получено этим способом, и к его семенам к способу получения трансформированного растения батата с высоким накоплением антоциана, в котором растение имеет повышенную устойчивость к стрессу, обусловленному окружающей средой, по сравнению с растением дикого типа благодаря трансформации растения батата, способного накапливать антоциан, рекомбинантным вектором к трансгенному растению батата с высоким накоплением антоциана и повышенной устойчивостью к стрессу,обусловленному окружающей средой, по сравнению с растением дикого типа, где трансгенное растение получено с помощью вышеупомянутого способа и к генной вставке для повышения содержания каротиноида и повышения устойчивости к стрессу,обусловленному окружающей средой, растения батата, способного накапливать антоциан, которое содержит вышеупомянутую - модификацию гена.(73) КОРЕА РИСРЧ ИНСТИТЬЮТ ОФ БАЙОСАЙЕНС ЭНД БАЙОТЕКНОЛОДЖИ(74) Русакова Нина Васильевна Жукова Галина Алексеевна Ляджин Владимир Алексеевич Ляджин Алексей Владимирович Иванова Антонина Сергеевна(54) СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯС ВЫСОКИМ НАКОПЛЕНИЕМ КАРОТИНОИДА И АНТОЦИАНА И РАСТЕНИЕ, ПОЛУЧЕННОЕ ЭТИМ СПОСОБОМ(57) Настоящее изобретение относится к способу получения растения батата с высоким накоплением ОБЛАСТЬ ПРИМЕНЕНИЯ Настоящее изобретение относится к способу получения растения батата с высоким накоплением каротиноида и антоциана и к растению,полученному с помощью этого способа. Точнее сказать, оно относится к способу получения растения с высоким накоплением каротиноида и антоциана путем трансформации растения батата,способного накапливать антоциан, с помощью рекомбинантного вектора, содержащего модификацию гена, имеющую нуклеотидную последовательность, искусственно вставленную в ген , выделенный изк трансформированному растению батата с высоким накоплением каротиноида и антоциана, которое получено этим способом, и к его семенам к способу получения трансгенного растения батата с высоким накоплением антоциана, в котором растение имеет повышенную по сравнению с растением дикого типа устойчивость к стрессу,обусловленному окружающей средой, благодаря трансформации растения батата, способного накапливать антоциан,с помощью рекомбинантного вектора к трансгенному растению батата с высоким накоплением антоциана, которое получено с помощью вышеупомянутого способа и имеет повышенную по сравнению с растением дикого типа устойчивость к стрессу,обусловленному окружающей средой, и его семенам к генной вставке для повышения содержания каротиноида и устойчивости к стрессу,обусловленному окружающей средой, растения батата, способного накапливать антоциан,которое содержит вышеупомянутую модификацию гена -. ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ Батат (.) представляет собой корнеплод, который может культивироваться на относительно бедной почве с урожайностью около 30 тонн с гектара, он употребляется в качестве пищи, а также пищевого продукта для скота. Окрашенный батат, имеющий фиолетовый цвет, желтый цвет или подобный им, содержит различные антиоксиданты. В частности, желтый батат, батат желтого цвета, содержит бета-каротин в количестве 14,7-20 мг/100 г, а фиолетовый батат содержит антоциан в количестве 2,28 г/100 г, чем обусловлена их превосходная антиоксидантная активность, обеспечивающая удаление активного кислорода, который содействует старению клеток и является причиной различных заболеваний у взрослых. С 1990-х годов была опробована трансформация путем сокультивирования си разработана система регенерации и трансформации растений путем индукции культуральных клеток с эмбриогенной компетенцией и эмбриогенеза соматических клеток из апикальной и латеральной меристем (.19, 227-239, 2007). Однако о батате, способном продуцировать высокий уровень низкомолекулярных антиоксидантов, таких как каротиноид, антоциан и полифенол, публикаций еще не имеется. Таким образом, получение новых сельскохозяйственных культур,имеющих 2 устойчивость к разнообразным стрессам и способных продуцировать высокий уровень низкомолекулярных антиоксидантов, может дать свой вклад в решение проблем, относящихся к продовольствию, энергии и окружающей среде, с которыми столкнется человечество в 21 столетии.и другие нашли, что в листьях батата присутствуют по крайней мере 15 видов антоциана,обладающих высокой физиологической активностью (. 2002 . .. 83, 1076-1082). Интересно, что, как было отмечено в публикации, биосинтез антоциана в листьях многих растений индуцирован экстремальным стрессом, обусловленным внешней средой, который вызывает генерацию активного кислорода, известно также, что антоциан в листьях или, в особенности, в корнях играет важную роль в механизме защиты растения от экстремальных воздействий (. 2002. 25,539-547). Общеизвестно, что фенольные соединения растений, благодаря их многообразию и широкой распространенности, являются наиболее важными естественными антиоксидантами. Полифенол и фенольные соединения также привлекают к себе внимание, так как известно, что они играют важную роль в защите человеческого организма против различных окислительных стрессов. Гидроксикоричные кислоты (НСА) являются представителями полифенолов, найденные в корнях батата и известные как сильные антиоксиданты, они содержатся в большом количестве в большинстве видов батата независимо от вида тканей или времени развития растения батата (. 2002. .. 50, 3718-3722). Таким образом,так же как каротиноид и антоциан, полифенол вносит большой вклад в проявление высокой антиоксидантной активности батата. Всемирная организация здравоохранения (ВОЗ) сообщает, что более ста миллионов детей в мире страдают от дефицита витамина А, и каждый год более половины миллиона детей теряют зрение из-за этого дефицита. Бета-каротин как предшественник витамина А обладает физиологической активностью в качестве пищевой и биологически активной добавки и, таким образом, исследования по искусственному изменению метаболизма,касающиеся накопления каротиноида в продуктах питания, являются очень важными для улучшения их пищевой ценности. Известно, что бета-каротин обладает физиологической активностью как вещество с высокими антиоксидантными свойствами, и он также играет важную роль в собственном механизме защиты растений от оксидативного стресса. Недавно было обнаружено,что на биосинтез каротиноида оказывает влияние абсцизовая кислота (АВА), которая является одним из видов растительных гормонов, связанных со стрессом, вызванным окружающей средой, поэтому возникла необходимость в исследованиях, 30683 касающихся взаимосвязи этих антиоксидантных соединений и стрессов,обусловленных окружающей средой. Несмотря на то, что в направлении получения полезных материалов из трансгенных растений проведено много исследований, коммерческое применение этих методов не достигло должного уровня из-за низкой продуктивности. В связи с этим считается, что способ снижения стоимости продукции путем использования культивированных растительных клеток или растений сам по себе будет более выгодным для экономически приемлемого массового производства продукции с высоким содержанием физиологически активного материала. Кроме того, если создано растение,способное производить полезные материалы в большом количестве, и из него получены культивируемые растительные клетки, время,требуемое для достижения производства полезных материалов в больших масштабах, будет сокращено и будет легко достигнуто высокое качество культуры выращиваемых клеток. Кроме того, может быть исключена проблема,связанная с экологическими рисками,которые могут встречаться при выращивании трансгенных растений, поэтому данный способ может широко применяться для получения различных целевых соединений, таких как физиологически активные протеины. Известно также, что, по сравнению с выращиванием микроорганизмов или животных клеток, применение этого метода менее негативно воспринимается общественностью, даже когда он применяется для производства фармацевтических или пищевых продуктов. Корейский патент, регистрационный номер. 10-0813284, раскрывает способ повышения содержания каротиноида путем трансформации растения геном фермента для биосинтеза фитоена,выделенным из мандарина, а корейская выложенная заявка . 2010-0100097 раскрывает способ повышения устойчивости растения к солевому стрессу путем трансформации растения геном 1, выделенным из батата. ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ Настоящее изобретение задумано под влиянием описанных выше обстоятельств, и авторы настоящего изобретения завершили настоящую работу созданием растения батата с высоким накоплением каротиноида и антоциана путем трансформации растения батата, обладающего способностью накапливать антоциан,рекомбинантным вектором, содержащим модификацию гена, имеющую специфическую нуклеотидную последовательность, искусственно вставленную в ген , выделенный из, а также подтверждением проявления трансгенным растением устойчивости к стрессу,обусловленному окружающей средой. ТЕХНИЧЕСКИЕ СРЕДСТВА ДЛЯ РЕШЕНИЯ ПРОБЛЕМ Для решения проблем, описанных выше, в настоящем изобретении заявляется способ получения растения батата с высоким накоплением каротиноида и антоциана,включающий трансформацию растения батата, способного накапливать антоциан, рекомбинантным вектором,содержащим - модификацию гена, имеющую специфическую нуклеотидную последовательность,искусственно вставленную в ген ,выделенный из, и повышенную экспрессию - модификации гена. Заявляется также трансгенное растение батата с высоким накоплением каротиноида и антоциана,полученное данным способом, и его семена. Заявляется также способ получения растения батата с высоким накоплением антоциана и повышенной устойчивостью к стрессу,обусловленному окружающей средой, по сравнению с растением дикого типа,включающий трансформацию растения батата, способного накапливать антоциан, рекомбинантным вектором,включающим модификацию гена,имеющую специфическую нуклеотидную последовательность, искусственно вставленную в ген , выделенный из, и повышенную экспрессию - модификации гена. Заявляется также трансформированное растение батата с высоким накоплением антоциана, которое получают с помощью вышеупомянутого способа и которое имеет повышенную устойчивость к стрессу,вызванному окружающей средой, по сравнению с растением дикого типа. Заявляется также генная вставка для повышения содержания каротиноида в растении батата,способном накапливать антоциан, где генная вставка содержит - модификацию гена,имеющую специфическую нуклеотидную последовательность, искусственно вставленную в ген , полученный из. Заявляется также генная вставка для повышения устойчивости к стрессу, вызванному окружающей средой, растения батата, способного накапливать антоциан, где генная вставка содержит модификацию гена, имеющую специфическую последовательность, искусственно вставленную в ген , полученный из. ПОЛЕЗНЫЙ ЭФФЕКТ ИЗОБРЕТЕНИЯ Согласно настоящему изобретению, показано,что трансгенный батат с высоким накоплением каротиноида и антоциана,полученный в соответствии с настоящим изобретением, может быть использован как функциональный пищевой продукт, который также, благодаря проявлению устойчивости к стрессу, вызванному окружающей средой, будет весьма полезен для повышения урожайности сельскохозяйственных культур. КРАТКОЕ ОПИСАНИЕ РИСУНКОВ Фиг.1 иллюстрирует растительный вектор экспрессии гена- модификации гена, полученного из батата,с использованием соматического эмбриогенеза 3 для трансгенного растения (/ (- трансгенное растение). Фиг.3 иллюстрирует (А) поперечный срез запасающего корня трансгенного батата сортасо сверхэкспрессией - модификации гена, полученного из батата по настоящему изобретению,и(В) наземную часть трансформированного растения ( нетрансгенные растения,/ контрольное растение,трансформированное пустым вектором, и трансгенное растение,/ растение,растение, с стебель / растениястебель растения, е). Фиг.4 иллюстрирует профиль экспрессии генов,связанных с биосинтезом каротиноида,в трансгенном растении , которое было трансформировано - модификацией гена,полученного из батата, где профиль экспрессии гена был определен с помощью - ( с обратной транскрипцией) и электрофореза. Фиг.5 иллюстрирует профиль экспрессии генов,участвующих в биосинтезе антоциана,в трансгенном растении , которое было трансформировано - модификацией гена,полученного из батата, где профиль экспрессии генов был определен с помощью - и электрофореза. Фиг.6 иллюстрирует результат измерения полного содержания каротиноида в запасающих корнях трансгенного растениясо сверхэкспрессией - модификации гена,полученного из батата, где измерения были выполнены с использованием спектрофотометра. Фиг.7 иллюстрирует результат измерения полного содержания антоциана в запасающих корнях трансгенного растениясо сверхэкспрессией - модификации гена,полученного из батата, где измерения были выполнены с использованием спектрофотометра. Фиг.8 иллюстрирует результат анализа(2,2-дифенил-1-пикрилгидразил) активности захвата свободных радикалов и(В) содержания полифенолов в листьях трансгенного растениясо сверхэкспрессией - модификации гена, полученного из батата. Фиг.9 иллюстрирует результат анализа устойчивости к -стрессу при различных концентрациях соли в листовом диске трансгенного растениясо сверхэкспрессией модификации гена, полученного из батата. Фиг.10 иллюстрирует результат анализа устойчивости к стрессу при -обработке( метилвиологен) при различных концентрациях в листовом диске трансгенного растениясо сверхэкспрессией модификации гена, полученного из батата. 4 ЛУЧШИЙ(ИЕ) РЕЖИМ(Ы) ДЛЯ ВЫПОЛНЕНИЯ ИЗОБРЕТЕНИЯ. Для достижения цели настоящего изобретения,описанной выше, настоящее изобретение предлагает способ получения растения батата с высоким накоплением каротиноида и антоциана,включающий трансформацию растения батата,способного накапливать антоциан, рекомбинантным вектором, содержащим - модификацию гена,состоящую из нуклеотидной последовательностиО 1 и содержащую специфическую нуклеотидную последовательность, искусственно вставленную в ген , полученный из, со сверхэкспрессией - модификации гена. Согласно настоящему изобретению, модификация гена была получена путем встраивания аминокислотного участкав ген(см. Корейский патент,регистрационный номер . 0990330). Однако, хотя эта модификация, служит для повышения содержания каротиноида или повышения солеустойчивости, только этим она не ограничена. Предпочтительно, чтобы - модификация гена по настоящему изобретению могла содержать нуклеотидную последовательность,которая представлена последовательностью 1. Далее, в область охвата настоящего изобретения включены также модификации вышеупомянутой нуклеотидной последовательности. В частности,упомянутый выше ген может содержать нуклеотидную последовательность, предпочтительно имеющую по крайней мере 70, более предпочтительно - по крайней мере 80, еще более предпочтительно - по крайней мере 90, и наиболее предпочтительно - по крайней мере 95 гомологию с нуклеотидной последовательностью 1.гомологии последовательности определенного полинуклеотида определяется путем сопоставления сравниваемых участков двух последовательностей,которые выравнены оптимальным образом. В этом отношении часть полинуклеотида в сравниваемом участке может содержать вставки или делеции (т.е. промежутки) по сравнению с последовательностьюобразцом (не содержащим каких-либо вставок или делеций) при оптимизированном выравнивании двух последовательностей. Термин рекомбинантная обозначает клетку,которая реплицирует гетерогенные нуклеотиды или экспрессирует указанные нуклеотиды, или пептид,или гетерогенный пептид,или протеин,закодированный гетерогенными нуклеотидами. Рекомбинантная клетка может экспрессировать ген или фрагмент гена в виде смысловой или антисмысловой последовательности, которые не встречаются в естественном состоянии клетки. Кроме того, рекомбинантная клетка может экспрессировать ген, который встречается в естественном состоянии, после того как упомянутый ген модифицируется и вводится вновь в клетку искусственным образом. Согласно настоящему изобретению,последовательность - модификации гена может быть включена в рекомбинантный вектор экспрессии. Термин рекомбинантный вектор экспрессии означает бактериальную плазмиду, фаг,дрожжевую плазмиду, вирус растительной клетки,вирус клетки млекопитающих или другой вектор. Обычно может быть использована любая плазмида и вектор, если она/он может размножаться или устойчиво существовать в хозяине. Важной характеристикой вектора экспрессии является то,что он включает начало репликации, промотор,репортерный ген и элемент контроля трансляции. Вектор экспрессии включает последовательность ДНК для кодирования - - протеина и подходящие элементы контроля транскрипции/трансляции, которые могут быть сконструированы с помощью способа, хорошо известного специалистам в данной области. Данный способ включает метод рекомбинантной ДНК, метод синтеза ДНК и метод рекомбинантной технологии. Для индукции синтеза мРНК последовательность ДНК может быть эффективно скомпонована с соответствующим промотором,имеющимся в векторе экспрессии. Кроме того,вектор экспрессии может включать сайт для связывания с рибосомой в качестве сайта инициации трансляции, а также терминатор транскрипции. Предпочтительным примером рекомбинантного вектора согласно настоящему изобретению является-плазмидный вектор, который может передавать часть себя, т.е. так называемую Т-область, в растительную клетку, когда данный вектор присутствует в подходящей клетке-хозяине, такой как. Другие типы-плазмидного вектора (см. ЕР 0 116 718 В 1) используются в настоящее время для передачи гибридной ДНК-последовательности в протопласты,что может привести к образованию нового растения путем соответствующего встраивания ДНК растительной клетки или гибридной ДНК в геном растения. Особенно предпочтительной формой-плазмидного вектора является так называемый бинарный вектор, раскрытый в ЕР 0 120 516 В 1 и патенте США . 4,940,838. Другие векторы,которые могут быть использованы для введения ДНК по настоящему изобретению в растениехозяина, могут быть выбраны из двуспирального растительного вируса),односпирального вируса и вирусного вектора,который может происходить из вируса , и т.д., например, неполный растительный вирусный вектор. Использование указанного вектора может быть особенно полезным в случаях, когда растениехозяин плохо трансформируется. Вектор экспрессии может включать по крайней мере один селективный маркер. Упомянутый селективный маркер представляет собой нуклеотидную последовательность, которая может быть выбрана путем использования обычных химических методов. Примеры включают все гены,которые пригодны для того, чтобы отличить трансформированные клетки от нетрансформированных. Конкретные их примеры включают ген устойчивости к гербицидам, таким как глифосат и фосфинотрицин, и ген устойчивости к антибиотикам, таким как канамицин, 418,блеомицин, гигромицин и хлорамфеникол, но не ограничиваются ими. Для рекомбинантного вектора согласно настоящему изобретению промотор может быть любым из 35, актина, убиквитина, , или гистоновым промотором, но не ограничивается ими. Термин промотор означает молекулу ДНК, с которой связывается РНКполимераза, чтобы инициировать ее транскрипцию,и она соответствует участку ДНК перед структурным геном. Термин растительный промотор означает промотор, который может инициировать транскрипцию в растительной клетке. Термин конститутивный промотор означает промотор, который активен при большинстве условий окружающей среды и на большинстве стадий развития или стадий дифференцировки клеток. Так как может быть выбран трансформант с различными механизмами на различных стадиях,конститутивный промотор может быть предпочтительным для настоящего изобретения. Поэтому возможность для выбора конститутивного промотора здесь не ограничивается. Для рекомбинантного вектора по настоящему изобретению может использоваться любой общеиспользуемый терминатор. Примеры включают терминатор нопалин синтазы , терминатор а-амилазы 1 риса, терминатор гена фазеолина, терминатор гена октопинаи т.д., но не ограничиваются ими. В отношении необходимости терминатора в основном известно, что наличие такой области может повышать надежность и эффективность транскрипции в растительных клетках. Поэтому использование терминатора крайне предпочтительно с точки зрения содержания настоящего изобретения. Трансформация растения означает любой метод,с помощью которого ДНК доставляется в растение. Такой метод трансформации не требует времени для регенерации и/или использования культуры ткани. Трансформация растительных видов в настоящее время довольно распространена не только для двудольных растений, но также и для однодольных. В принципе, для введения гибридной ДНК по настоящему изобретению в соответствующие исходные клетки может быть использован любой способ. Способ может быть соответственно выбран из метода слияния протопластов, индуцированного кальцием/полиэтиленгликолем (, .,1982,296,72-74..,1987, 8, 363-373), метода электропорации для протопластов ( ., 1985/. 3, 1099-1102), метода введения с помощью микроинъекций для растительных компонент ( . . ., 1986, . 202, 179-185), метода бомбардирования частицами(которые покрыты ДНК или РНК) для различных растительных компонент ( Т.М.., 1987, 327, 70) или метода неполной вирусной инфекции для переноса гена с помощью 5 путем заражения растения или трансформации полностью созревшей пыльцы или микроспор (ЕР 0 301 316) и т.д. Предпочтительным способом в настоящем изобретении является перенос ДНК с помощью. В частности, для настоящего изобретения предпочтительным является использование так называемой бинарно-векторной технологии, раскрытой в ЕР А 120 516 и в. 4,940,838. Согласно способу по одному воплощению настоящего изобретения, бататом, способным накапливать антоциан, предпочтительно является батат фиолетового цвета, и более предпочтительно батат сорта , но не ограничивается ими. В настоящем изобретении заявляется также трансгенное растение батата с высоким накоплением каротиноида и антоциана, полученное этим методом, и его семена. По сравнению с нетрансформированным растением,батат сорта,трансформированный - модификацией гена,полученного из батата по настоящему изобретению,демонстрирует повышенную экспрессию гена, а также , -, - и -,которые являются основными генами,относящимися к биосинтезу каротиноида, и было также подтверждено, что содержание каротиноида возросло примерно в 3 раза или более по сравнению с нетрансформированным растением (см. фиг.4 и фиг.6). В настоящем изобретении заявляется также способ получения растения батата с высоким накоплением антоциана и повышенной устойчивостью к стрессу,обусловленному окружающей средой, включающий трансформацию растения батата, способного накапливать антоциан,рекомбинантным вектором, содержащим модификацию гена, состоящую из нуклеотидной последовательностиО 1 и имеющую специфическую нуклеотидную последовательность,искусственно вставленную в ген ,полученный из, со сверхэкспрессией- модификации гена. Согласно способу одного из воплощений настоящего изобретения, растением, способным накапливать антоциан, предпочтительно является фиолетово окрашенный батат,и более предпочтительно - батат сорта , но не ограничивается ими. Согласно одному из воплощений настоящего изобретения стрессом, обусловленным окружающей средой, может быть абиотический стресс и более предпочтительно - солевой или метилвиологенный стресс, но не ограничивается ими. В настоящем изобретении заявляется также трансформированное растение батат с высоким по сравнению с диким типом накоплением антоциана и повышенной устойчивостью к стрессу,обусловленному окружающей средой, которое получено предлагаемым способом, и его семена. В настоящем изобретении заявляется также генная вставка для повышения содержания 6 каротиноида в растении батата, способном накапливать антоциан, в котором генная вставка содержит - модификацию гена, состоящую из нуклеотидной последовательностиО 1 и содержащую специфическую нуклеотидную последовательность, искусственно вставленную в ген , полученный из. Генная вставка по настоящему изобретению содержит в качестве действующей компоненты модификацию гена, состоящую из нуклеотидной последовательностиО 1, и путем трансформации этой модификацией гена растения батата,способного накапливать антоциан,становится возможным повысить содержание каротиноида в растении батата, способном накапливать антоциан. В настоящем изобретении заявляется также генная вставка для повышения устойчивости к стрессу, обусловленному окружающей средой,растения батата, способного накапливать антоциан,где генная вставка содержит - модификацию гена,состоящую из нуклеотидной последовательности 1 и содержащую специфическую нуклеотидную последовательность,искусственно вставленную в ген ,полученный из. Генная вставка по настоящему изобретению содержит в качестве действующей компоненты - модификацию гена,состоящую из нуклеотидной последовательностиО 1, и путем трансформации этой модификацией гена растения батата,способного накапливать антоциан,становится возможным повысить устойчивость растения батата, способного накапливать антоциан,к стрессу, обусловленному окружающей средой. Ниже приводится более подробное пояснение настоящего изобретения на Примерах. Однако очевидно, что приведенные Примеры даны только для иллюстрации настоящего изобретения, и оно никоим образом не ограничивается ими. ПРИМЕРЫ Пример 1 Клонирование и анализ нуклеотидной последовательности - - гена. Были приготовлены-праймеры для клонирования-гена из. Праймеры имеют следующие последовательности прямой праймер (532) и обратный праймер(5-33). Используя полученныйген в качестве шаблона,была выполнена перекрывающаясядля того,чтобы иметь искусственную мутацию специфической нуклеотидной последовательности в. Для вставки последовательности оснований,отвечающей последовательности(см. Корейский патент, регистрационный номер . 0990330), были приготовлены -праймеры, и последовательности в использованных праймерах были следующие прямой праймер-продукт размером в 921 пару нуклеотидов, и он был клонирован в - - вектор. Наличие мутации - вставки последовательности ,начинающейся с аминокислоты номер 133, было подтверждено путем выполнения секвенирования. была выполнена с использованием полимеразной смеси 2 фирмы , и-продукт желаемых размеров был клонирован с использованием системы и затем секвенирован для определения полной нуклеотидной последовательности. Полученный в результате кДНК ген был назван-. Что касается полной длины - гена по настоящему изобретению, то эта кДНК из 924 пар нуклеотидов кодирует 307 аминокислот. Изоэлектрическая точкаи молекулярный вес,которые были установлены из аминокислотной последовательности, составили соответственно 8,45 и 33,74 к. К нуклеотидной последовательности описанных выше праймеров, к 5 концу каждого описанного выше праймера,были добавлены последовательности-адаптеры (большие буквы) так,чтобы можно было использовать технологию клонирования , производимую фирмой. Получены следующие нуклеотидные последовательности прямой праймер(5 С-37). После клонирования -продукт,имеющий желаемые размеры, клонировался с помощью системы -,полная нуклеотидная последовательность была проверена путем секвенирования. Полученная кДНК была названа -. Каждый из двухвекторов,в которые были скопированы соответственно генили ген -, были подвергнуты ВР-реакции, чтобы клонировать гены в 207 векторе. После этого был сконструирован растительный вектор экспрессии геновили -. Эти гены были клонированы в соответствии среакцией между 207 и 11 векторами, в результате чего был сконструирован растительный вектор экспрессии (фиг.1). Пример 2 Трансформация культивированных соматических клеток бататамодификацией гена, полученного из батата. Культивированные соматические клетки батата, которые представляли собой поколение,имеющее высокий уровень антоциана, были трансформированы - модификацией гена,полученного из батата под управлением 35 промотора (фиг.2 А). В результате было получено мини трансгенное растение,при этом использовалась селективная среда и проверка с помощьюгеномной ДНКэкспрессии гена гигромицина , который является селективным маркером для трансформанта, по которому производился отбор трансформанта. Полученный в результате этого трансформант был обозначен как - (фиг.2 В). Пример 3. Анализ фенотипа батата ,трансформированного - модификацией гена,полученного из батата. После выполнения трансформации ,который представляет собой фиолетово окрашенный батат, - модификацией гена,полученного из батата, и получения его запасающих корней было обнаружено, что в то время как нетрансгенныйдемонстрировал без изменения сильный фиолетовый цвет, как показано на фиг.3, - демонстрировал фенотип, в котором смешивались желтая и фиолетовая окраски(фиг.3 А). Кроме того, в фенотипе стебля было обнаружено, что он имеет многочисленные развитые боковые отростки и боковые ветви, а также большое количество листьев. (фиг.3 В). Пример 4. Анализ экспрессии генов,относящихся к биосинтезу каротиноида, в батате,трансформированном модификацией гена из батата. Для нетрансгенного и трансгенного бататаэкспрессия генов, относящихся к биосинтезу каротиноида, и экспрессия генаанализировались на основе -. В результате, в случае нетрансгенного растения и растения,трансформированного пустым вектором, экспрессия гена была либо слабой,либо отсутствовала, тогда как в - экспрессия генасущественно возрастала. Кроме того, в случае нетрансгенного растения и растения,трансформированного пустым вектором, экспрессия генов, относящихся к биосинтезу каротиноида,была, как правило, либо слабой, либо отсутствовала,тогда как экспрессия основных генов, относящихся к биосинтезу каротиноидов, таких как , -,- или -, в - была повышенной. Кроме того, повышалась также экспрессия генов,относящихся к биосинтезу стриголактона, таких как ген 4 или ген , и экспрессия гена,относящегося к биосинтезу АВА, такого как ген(фиг.4). Пример 5. Анализ экспрессии генов,относящихся к биосинтезу антоциана, в батате-. В результате,в случае нетрансформированного растения бататаи растения батата , трансформированного пустым вектором, экспрессия генов, относящихся к биосинтезу антоциана, была очень сильной, тогда как экспрессия основных генов, относящихся к биосинтезу антоциана, таких как , , 3, или 3, в - была снижена. Однако, в случае гена, относящегося к сосудистому накоплению антоциана, такого как 24, не наблюдалось существенного отличия в экспрессии по сравнению с нетрансгенным растением (фиг.5). 7 Пример 6. Анализ содержания каротиноида в батате , трансформированном модификацией гена, полученного из батата. Для того, чтобы увидеть изменения в содержании каротиноида в трансгенном батате, с помощью спектрофотометра было измерено содержание каротиноида в запасающих корнях нетрансгенного и трансгенного бататапутем измерения поглощения на 440 нм. В результате было получено, что трансгенный бататдемонстрирует возрастание содержания бета-каротина почти в 3 раза или более по сравнению с нетрансгенным растением, служащим в качестве контрольной группы (фиг.6). Пример 7. Анализ содержания антоциана в батате , трансформированном модификацией гена, полученного из батата. Для того, чтобы увидеть изменение в содержании антоциана в трансгенном батате ,проводилось измерение с помощью спекрофотометра содержания антоциана в запасающих корнях нетрансгенного и трансгенного батата . В результате было получено, что трасгенный батат демонстрировал снижение содержания антоциана примерно в 0,6 0,8 раза или более по сравнению с нетрансгенным растением, взятым в качестве контрольной группы(фиг.7). Однако в случае трансгенного растения 1-1 содержание антоциана было почти такое же, как и у нетрансгенного растения, и в этой трансгенной линии содержание каротиноида было наивысшим, и,таким образом, оно было выбрано в качестве идеального трансгенного растения (фиг.6 и фиг.7). Пример 8. Анализ материала с низким молекулярным весом, имеющего антиоксидантную активность,в листьях батата,трансформированного - модификацией гена,полученного из батата. Для сравнения содержания материала с низким молекулярным весом,обладающего антиоксидантной активностью,в листьях трансгенного бататабыла измеренаактивность захвата радикалов (Фиг.8 А). В результате было получено, что нетрансгенное растение или растение, трансформированное пустым вектором, демонстрировали активность,соответствующую примерно 250-340 мкг аскорбиновой кислотына грамм сырого веса( -), а трансгенное растение демонстрировало активность, соответствующую примерно 310-610 мкг аскорбиновой кислоты. Трансгенное растение 1-1 демонстрировало практически наивысшую антиоксидантную активность. Кроме того, в отношении содержания полифенола нетрасгенное растение или растение,трансформированное пустым вектором,демонстрировали активность, соответствующую примерно 170-190 мкг хлорогеновой кислоты на грамм сырого веса , а трансгенное растение демонстрировало активность, соответствующую примерно 230-360 мкг хлорогенной кислоты,демонстрируя тем самым более высокое содержание полифенола в трансгенном растении по сравнению с контрольной группой (фиг.8 В). Пример 9. Анализ устойчивости к абиотическому стрессу батата , трансформированного,трансформированный модификацией гена, полученного из батата,демонстрировал возрастание бета каротина в 2,5 2,9 раза по сравнению с нетрансгенным . Поэтому, чтобы подтвердить, обусловлена ли устойчивость трансгенного растения к стрессу наличием антоциана и каротиноида, содержание которых увеличивается благодаря настоящему изобретению,растение было подвергнуто абиотическому стрессу, такому как солевой стресс и-стресс. Сначала из верхних четырех-пяти листьев с вершины листового стебля контрольной группы и трансгенного растения были подготовлены листовые диски диаметром 8 мм, а затем их напитывалив растворе с концентрацией 0, 150,200 или 400 мМ в течение 24 часов в условиях темноты при 25 С. После этого оксидативный стресс, испытываемый каждым листовым диском,наблюдали с помощью(3,3 диаминобензидин) окрашивания. При реакциис Н 2 О 2,присутствующей в клетке, образуется коричневый осадок. После 24 часового насыщения в растворе 1 мг/мл - ( 3,8) каждой испытываемой группы путем фотографирования регистрировалась степень окрашивания. В результате было обнаружено,что в случае,трансформированного пустым вектором, степень окрашивания в коричневый цвет становилась сильнее с ростом концентрации , указывая на то,что растение испытывало больший оксидативный стресс с ростом концентрации, тогда как трансгенное растение демонстрировало меньшую степень окрашивания по сравнению с контрольной группой, указывая на устойчивость к оксидативному стрессу, вызванному солью (Фиг.9). Для проверки на метил-виологеновыйстресс как пример оксидативного стресса,вызванного другой причиной, были выбраны шесть листовых дисков диаметром 8 мм, и каждый из них был помещен в многолуночный культуральный планшет с диаметром лунок 2,5 см с 3 мл 0,4 раствора сорбитола в каждой лунке, содержащего 0,2,5, 5 или 10 мкМ . После выдержки для поглощенияв течение 12 часов в темноте при 25 С культивирование продолжалось в течение 24 часов в условиях освещения. После этого измерялась ионная проводимость раствора с интервалом 12 часов, для этого использовали электрокондуктометр (модель 455 С, , .,Корея) и через 24 часа измеряли оксидативный стресс с помощьюокрашивания для определения степени повреждения листа. Нетрансгенное растение и растение контрольной группы, трансформированное пустым вектором,демонстрировали высокую ионную проводимость в 37,3 и 55,7 соответственно в течение времени до 48 часов после обработки 2,5 мкМ , что указывало на высокую чувствительность в отношении оксидативного стресса, вызванного . Трансгенное растение, напротив, демонстрировало низкую ионную проводимость в 25,6 и 31,2 соответственно, что указывало на высокую устойчивость к оксидативному стрессу, вызванному. Нетрансгенное растение и растение контрольной группы, трансформированное пустым вектором, демонстрировали высокую ионную проводимость в 51,9 и 62,5 соответственно в течение времени до 48 часов после обработки 5 мкМ, что указывало на высокую чувствительность по отношению к оксидативному стрессу, вызванному. Трансгенное растение,напротив,демонстрировало низкую ионную проводимость в 34,8 и 40,8 соответственно, что указывало на высокую устойчивость к оксидативному стрессу,вызванному(Фиг.10 А). Кроме того, в результате наличия окрашивания после пропитывания каждой обрабатываемой группы в течение 24 часов в растворе 1 мг/мл - ( 3,8) получено, что нетрансгенное растение и растение контрольной группы, трансформированное пустым вектором, демонстрировали более высокую степень окрашивания в коричневый цвет при росте концентрации , что указывало на то, что растение испытывает больший оксидативный стресс при росте концентрации. Трансгенное растение,напротив, демонстрировало более низкую степень окрашивания по сравнению с контрольной группой,что указывало на устойчивость к оксидативному стрессу, вызванному(фиг.10 В). ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Способ получения растения батата с высоким накоплением каротиноида и антоциана,включающий трансформацию растения батата,способного накапливать антоциан, рекомбинантным вектором, содержащим - модификацию гена,состоящую из нуклеотидной последовательности 01 и содержащую специфическую нуклеотидную последовательность, искусственно вставленную в ген , полученный изсо сверхэкспрессией - модификации гена. 2. Способ по п.1, характеризующийся тем, что растением батата является батат, имеющий фиолетовую окраску. 3. Растение батата с высоким накоплением каротиноида и антоциана, полученное по способу по п.1 или п.2. 4. Семена растения батата по п.3. 5. Способ получения растения батата с высоким накоплением антоциана и повышенной устойчивостью к стрессу,обусловленному окружающей средой, по сравнению с растением дикого типа, включающий трансформацию растения батата,способного накапливать антоциан,рекомбинантным вектором, включающим модификацию гена, состоящую из нуклеотидной последовательности 01 и содержащую специфическую нуклеотидную последовательность,искусственно вставленную в ген ,полученный из, со сверхэкспрессией- модификации гена. 6. Способ по п.5, характеризующийся тем, что растением батата является батат, имеющий фиолетовую окраску. 7. Способ по п.5, характеризующийся тем, что стрессом, обусловленным окружающей средой,является абиотический стресс. 8. Растение батата с повышенной устойчивостью к стрессу, обусловленному окружающей средой, по сравнению с растением дикого типа и высоким накоплением антоциана, полученное по способу по п.5 или п.7. 9. Семена растения батата по п.8. 10. Генная вставка для повышения содержания каротиноида в растении батата, способном накапливать антоциан, которая содержит модификацию гена, состоящую из нуклеотидной последовательности 01 и содержащую специфическую нуклеотидную последовательность,искусственно вставленную в ген ,полученный из. 11. Генная вставка для повышения устойчивости к стрессу, обусловленному окружающей средой,растения батата, способного накапливать антоциан,которая содержит - модификацию гена,состоящую из нуклеотидной последовательности 01 и содержащую специфическую нуклеотидную последовательность, искусственно вставленную в ген , полученный из
МПК / Метки
МПК: C12N 15/82, A01H 5/00, C12N 15/29
Метки: каротиноида, ipomoea, этим, способ, растение, batatas, получения, полученное, растения, способом, высоким, антоциана, накоплением, трансгенного
Код ссылки
<a href="https://kz.patents.su/14-30683-sposob-polucheniya-transgennogo-rasteniya-ipomoea-batatas-s-vysokim-nakopleniem-karotinoida-i-antociana-i-rastenie-poluchennoe-etim-sposobom.html" rel="bookmark" title="База патентов Казахстана">Способ получения трансгенного растения ipomoea batatas с высоким накоплением каротиноида и антоциана и растение, полученное этим способом</a>
Ibor-ins модификация гена, полученный из ipomoea batatas, рекомбинантный вектор,клетка-хозяин, трансформированная рекомбинантным вектором, способ получения трансформированного растения, трансформированное растение, имеющее повышенное содержание каротиноида, способ получения трансформированного двудольного растения, трансформированное двудольное растение, имеющее повышенную устойчивость к солевому стpессу, генная вставка для повышения содержания каротиноида в растении и устойчивости к солевому стрессу растения

Номер патента: 29184
Опубликовано: 17.11.2014
Авторы: ЛИ, Хенг-Сун, АН, Йонг Ок, КВОК, Санг-Су, КИМ, Сун ха, ЖОНГ, Жэ Чол
МПК: A01H 5/10, A01H 5/00, C12N 15/29...
Метки: гена, растения, полученный, batatas, имеющее, двудольного, солевому, повышенное, растении, трансформированная, устойчивости, ipomoea, трансформированного, содержания, содержание, получения, повышенную, стрессу, вектор,клетка-хозяин, двудольное, способ, повышения, вектором, ibor-ins, рекомбинантный, устойчивость, вставка, трансформированное, генная, модификация, стpессу, растение, рекомбинантным, каротиноида
Текст:
...едовательность, искусственно вставленную вген, полученный из, которая состоит из нуклеотидной последовательности 2 10. Генная вставка для повышения устойчивости к солевому стрессу растения, включающаяген, полученный из, который состоит из нуклеотидной последовательности 1 или - модификацию гена,имеющую специфическую нуклеотидную последовательность, искусственно вставленную вген, полученный из,которая состоит из нуклеотидной последовательности...
Генетическая конструкция, содержащая специфический для фазы старения промотор, содержащие ее вектор, клетка и трансгенное растение, получаемый из трансгенного растения материал и собранный с него лист, способ повышения уровня треонина в листьях растения и предназначенный для курения продукт, содержащий обогащенный треонином табак

Номер патента: 27382
Опубликовано: 16.09.2013
Авторы: ЛИЧ, Гвендолин, КОУТС, Стив, ДЖОНС, Луиз
МПК: C12N 15/52, C12N 15/05, C12N 15/00...
Метки: способ, конструкция, вектор, содержащий, обогащенный, старения, генетическая, клетка, листьях, него, уровня, получаемый, специфический, лист, материал, курения, трансгенного, содержащая, собранный, промотор, трансгенное, растения, фазы, треонина, продукт, повышения, растение, табак, треонином, предназначенный, содержащие
Текст:
...оящем изобретении генетическая конструкция,содержащая специфический для фазы старения промотор, функционально связанный с кодирующей последовательностью,которая кодирует полипептид,обладающий активностью нечувствительной к треонину аспартаткиназы.(74) Тагбергенова Алма Таишевна Тагбергенова Модангуль Маруповна Касабекова Найля Ертисовна(54) ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ,СОДЕРЖАЩАЯ СПЕЦИФИЧЕСКИЙ ДЛЯ ФАЗЫ СТАРЕНИЯ ПРОМОТОР,СОДЕРЖАЩИЕ ЕЕ ВЕКТОР, КЛЕТКА...
Генетическая конструкция, кодирующая фосфоенолпируваткарбоксикиназу (рск) и/или пируватортофосфатдикиназу (ppdk); содержащие ее рекомбинантный вектор, клетка и трансгенные растения; способы получения трансгенного растения; материал для размножения растения; способ повышения концентрации рск и/или ppdk в листе опытного растения; способ снижения концентрации азота в листьях тестируемого растения; способ повышения скорости роста опытного растения; собранный с растения лист с низким уровнем азота и полученный из него продукт для курения

Номер патента: 28061
Опубликовано: 15.01.2014
Авторы: ЛИСС, Анна Элизабет, ТАЙЛОР, Люси Элизабет, ХИББЕРД, Джулиан Майкл
МПК: A01H 5/12, A24B 15/00, A01H 5/00...
Метки: пируватортофосфатдикиназу, собранный, генетическая, уровнем, опытного, листьях, вектор, способ, трансгенные, продукт, повышения, рекомбинантный, размножения, ppdk, получения, клетка, тестируемого, трансгенного, него, материал, растения, рск, полученный, курения, азота, кодирующая, концентрации, фосфоенолпируваткарбоксикиназу, снижения, низким, способы, содержащие, роста, листе, скорости, лист, конструкция
Текст:
...скорости роста стареющих растений. В заявке описаны также собранные листья растений, такие как листья табака,трансформированные генетическими конструкциями,а также предназначенные для курения продукты,содержащие указанные собранные листья.(74) Тагбергенова Модангуль Маруповна Тагбергенова Алма Таишевна Касабекова Найля Ертисовна(РСК) И/ИЛИ ПИРУВАТОРТОФОСФАТДИКИНАЗУСОДЕРЖАЩИЕ ЕЕ РЕКОМБИНАНТНЫЙ ВЕКТОР,КЛЕТКА И ТРАНСГЕННЫЕ РАСТЕНИЯ СПОСОБЫ...
Способ изготовления взрывчатого вещества и взрывчатое вещество, полученное этим способом
Номер патента: 8184
Опубликовано: 15.11.1999
Авторы: Уолтер Б. ГРИНС, Олдрич МАЧАСИК, Росс П. КЛАРК, Гари Р. ИК
МПК: C06B 31/28, C06B 47/14, F42D 5/04...
Метки: взрывчатого, вещества, взрывчатое, полученное, способ, этим, изготовления, вещество, способом
Формула / Реферат:
Изобретение относится к способу и составу для утилизации отходов, включающих энергосодержащие материалы. Определенные заранее количества отходов, включающих энергосодержащие материалы, смешивают с промышленными взрывчатыми веществами, что ведет к участию энергосодержащих материалов в процессе детонации и позволяет, таким образом, утилизировать энергосодержащие материалы, которые в противном случае попали бы в отходы. Отходы в измельченной форме,...
Способ ингибирования экспрессии гена в растительной клетке-мишени, способ получения растения, в котором генная экспрессия может быть ингибирована в растительной ткани-мишени, способ получения мужского стерильного растения кукурузы

Номер патента: 8416
Опубликовано: 15.04.2002
Авторы: БЕЛЛ, Филип, Джон, ДРЭЙТОН, Пол, Ричард, БРАЙТ, Саймон, Уилльям, Джонатан, ГРИНЛЕНД, Эндрю, Джеймс
МПК: C12N 15/63, C12N 15/82, A01H 5/00...
Метки: ингибирована, экспрессии, быть, ингибирования, получения, растительной, котором, кукурузы, стерильного, растения, генная, гена, мужского, ткани-мишени, может, клетке-мишени, способ, экспрессия
Формула / Реферат:
Способ ингибирования экспрессии гена в мишенированной растительной ткани, который включает стабильную трансформацию вида растительной клетки, из которой может быть регенерировано целое растение с помощью генного конструкта, несущего тканеспецифичный или специфичный для развития промотор, который действует в соответствующих клетках соответствующей мишенированной растительной ткани, и деструктивный ген, кодирующий белок, который при...
Предыдущий патент: Система управления потоком воздуха внутри системы дозирования сельскохозяйственного продукта
Следующий патент: Хитоолигосахариды и способы для применения при стимулировании роста сои
Случайный патент: Способ возделывания гороха с внесением азотного удобрения